" Prima parte
Enzima care catalizează etapa a patra a ciclului krebs este α-ceto glutarat dehidrogenază; această enzimă este un complex enzimatic foarte asemănător cu piruvatul dehidrogenază. Ambele sunt compuse din 48-60 de proteine în care sunt recunoscute trei activități enzimatice diferite și au, de asemenea, aceiași cofactori enzimatici; sunt enzime foarte asemănătoare, deoarece acționează pe substraturi similare: atât piruvat, cât și l "α-keto glutarate, are α- cetoacizi. Mecanismul de acțiune al celor două complexe enzimatice este același.
Atacul pirofosfatului de tiamină asupra carbonilului (C = O) al „α-cetoglutarat, duce la decarboxilarea acestuia și se formează derivatul carboxihidroxi propilic. Odată cu transferul ulterior la lipoamidă, are loc un proces redox intern, din care se obține derivatul carboxi lipoamidic sau succinil lipoamida.
Sucinil lipoamida reacționează apoi cu coenzima A pentru a da succinil coenzima A (care continuă în ciclul krebs) și lipoamida redusă care este reoxidată de FAD: FADH2 care se formează este reoxidat de NAD + și se obține NADH. Prin urmare, în această etapă a avut loc a doua eliminare a unui carbon din scheletul carbonos, sub formă de dioxid de carbon.
O grupare acil legată de coenzima A se află într-o formă activată, adică are un conținut ridicat de energie: de aceea este posibilă exploatarea energiei succinil coenzimei A.

În etapa a cincea a ciclului krebs, succinil coenzima A este supusă acțiunii lui succinil tiokinază; s-au făcut două ipoteze despre modul său de acțiune: vom descrie doar una dintre cele două, deoarece este cea mai acreditată. Conform acestei ipoteze, succinil coenzima A este atacată de azotul unei histidine (Hys) a enzimei: se eliberează coenzima A și un aduct derivat din histidină se formează ca intermediar, adică enzima succinil (sau succinil-Hys ); un ortofosfat acționează asupra acestui intermediar, ducând la eliberarea de succinat și la formarea fosfoenzimei. Fosfoenzima, atacată de difuzat de guanozină (PIB), produce guasnosină trifosfat (GTP) și enzima este eliberată. Din punct de vedere energetic GTP = ATP: legătura care furnizează energie este aceeași la ambele specii (este legătura anhidridă dintre fosforil Β și fosforil γ). În unele cazuri, GTP este utilizat ca material cu un conținut ridicat de energie, dar, de obicei, GTP este transformat în ATP prin acțiunea enzimei nucleozid difosfo kinază (NDPK); este o enzimă care se găsește în celule și catalizează următoarea reacție:
N1TP + N2DP → N1DP + N2TP
Generic NiTP ® Nucleoside Triphosphate
Generic NiDP ® nucleosid difosfat
Este o reacție reversibilă; în cazul nostru se întâmplă:
GTP + ADP → PIB + ATP
prin urmare, poate continua spre dreapta sau spre stânga chiar și pentru mici variații ale concentrațiilor reactivilor.
Dacă ciclul krebs se desfășoară la o viteză atât de mare încât să conducă la o producție de ATP mai mare decât necesarul de energie, există o disponibilitate redusă a ADP în timp ce există o mulțime de ATP: reacția catalizată de nucleozida difosfo-kinază este, atunci, direcționată spre stânga (GTP se acumulează dacă nucleozidul difosfo kinază nu are suficient substrat, adică ADP). Prin urmare, GTP este un semnal al disponibilității energiei și, prin urmare, încetinește ciclul krebs.
A șasea etapă a ciclului krebs duce la formarea fumaratului prin acțiunea lui succinat dehidrogenază; această enzimă dă o reacție stereospecifică, deoarece se formează întotdeauna trans nesaturat (este o alchenă), adică fumarat (în timp ce izomerul cis este maleatul). Succinat dehidrogenază se găsește pe membrana mitocondrială internă, în timp ce toate celelalte enzime ale ciclului krebs sunt împrăștiate prin mitocondrie.
Succinat dehidrogenaza are FAD ca cofactor; este inhibat de oxaloacetat (inhibare feed-back) în timp ce are succinat și fumarat ca modulator pozitiv (activator). ciclul krebs necesită energie, astfel încât singura posibilitate de a obține oxaloacetat de la pacient este că concentrația pacientului este foarte mare: malatul este unul dintre metaboliții cu cea mai mare concentrație în celule. Reacția care transformă malatul în oxaloacetat este, de asemenea, favorizată de faptul că concentrația de oxaloacetat este menținută scăzută prin acțiunea citratului sintază. Reacția catalizată de succinat dehidrogenază este, atunci, o reacție de autoalimentare și aceasta este singura modalitate de a face ca transformarea malatului în oxaloacetat să aibă loc.
Concentrația malatului mitocondrial trebuie să fie compatibilă cu concentrația malatului citoplasmatic: numai atunci când concentrația malatului mitocondrial este atât de mare încât să garanteze conversia malatului în oxaloacetat (în ciclul krebs), atunci malatul poate fi utilizat și în alte modalități (care sunt citoplasmatice): în citoplasmă malatul poate fi transformat în oxaloacetat din care aspartatul poate fi obținut prin acțiunea GOT (este o transaminază) sau glucoză prin gluconeogeneză.
Ne întoarcem la a șaptea etapă a ciclului krebs, acesta fiind catalizat de enzimă fumarasi: se adaugă apă într-un mod stereospecific pentru a produce L-malat.
În ultima etapă a ciclului Krebs, despre care am vorbit deja, acțiunea malat dehidrogenază. Această enzimă folosește o moleculă NAD + pentru acțiunea sa catalitică.
Am concluzionat astfel descrierea diferitelor etape ale ciclului krebs.
Ciclul krebs este total reversibil.
Pentru a crește viteza ciclului krebs, concentrația metaboliților prezenți în acel ciclu poate fi crescută; una dintre strategiile de creștere a vitezei ciclului krebs constă în transformarea unei părți a piruvatului care intră în mitocondrii în oxaloacetat (prin acțiunea piruvatului carboxilazei) și nu transformarea acestuia în acetil coenzima A: crește astfel concentrația de oxaloacetat este un metabolit al ciclului krebs și, prin urmare, crește viteza întregului ciclu.
În ciclul krebs trei NAD + sunt transformate în trei NADH și un FAD în FADH2 și, în plus, se obține un GTP: prin canalizarea puterii de reducere obținute din ciclul krebs, se produce ATP suplimentar; în lanțul respirator, puterea reducătoare este transferată de la NADH și FADH2 la oxigen: acest transfer se datorează unei serii de enzime situate pe membrana mitocondrială care, în acțiunea lor, conduc la producerea de ATP.
Procesele lanțului respirator sunt procese exergonice și energia eliberată este utilizată pentru a produce ATP; scopul celulei este de a exploata procesele exergonice pentru a face sinteza ATP. Pentru fiecare moleculă de NADH care intră în lanțul respirator se obțin 2,5 molecule de ATP și pentru fiecare FADH2 se obțin 1,5 molecule de ATP; această diversitate se datorează faptului că FADH2 intră în lanțul respirator la un nivel mai scăzut decât NADH.
Cu puterea reducătoare a metabolismului aerob, se obțin 30-32 ATP (219-233 kcal / mol) cu o eficiență de aproximativ 33% (eficiența metabolismului anaerob este de aproximativ 2%).
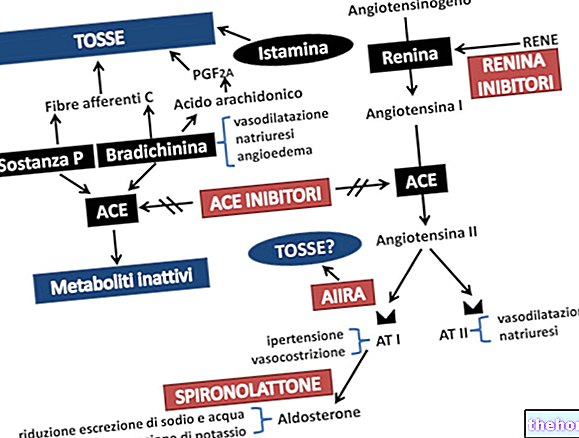







-cos-cause-sintomi-e-terapia.jpg)