ASOCIAȚIE (SAU „CONCATENARE” SAU „LEGĂTURĂ”)
Până în prezent am tratat încrucișarea dihidridă (sau polihibridă) mendeliană presupunând că diferitele perechi alelice se găsesc de fapt pe diferite perechi de cromozomi omologi. Dar numărul de perechi cromozomiale, deși diferă de la specie la specie, variază în limite înguste (puține specii ajunge la aproximativ o sută de cromozomi), în timp ce numărul de gene poate fi numărat în zeci de mii.
Faptul că personajele alese de Mendel pentru experimentele sale le-au separat separat (fără a confunda deci calculele în distribuția fenotipurilor în F2 a crucii polihibrice) a fost o șansă norocoasă. Dacă s-au găsit două perechi de alele pe loci adiacenți, legea care ar urma ar fi numit legea asocierii.
Știind că foarte multe caractere își au locusul pe un singur cromozom și că sunt perechile cromozomiale care se separă independent în meioză, se poate înțelege cât de des se întâmplă ca două perechi de caractere, dacă ar fi asociate pe un cromozom în părinți organism, rămân asociate în mod egal și în gamet și, prin urmare, în organismul căruia îi va aduce propriul său material genetic.
Astfel vedem că „asocierea reprezintă o„ excepție, departe de a fi rară, de la independența enunțată în a treia lege a lui Mendel.
SCHIMB SAU „CROSSING-OVER” ȘI RECOMBINARE
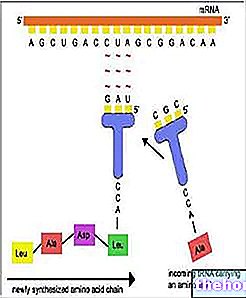
Vorbind despre meioză, am indicat că există două momente diferite de amestecare a materialului genetic: unul este cel al segregării cromozomilor în gamete și acesta este cel observat de Mendel.
Celălalt moment, care precede de fapt, este cel în care cele patru cromatide ale fiecărei perechi de cromozomi omologi schimb reciproc trăsături identice. În urma acestui schimb, doi factori care au fost asociați pe același cromozom vor fi în schimb independenți în gameți. faptul că are loc un schimb este proporțional, cu o primă aproximare, cu lungimea cromozomului, iar în cromozomii mai lungi poate exista chiar mai mult decât un schimb.
Fenomenul poate fi detectat citologic, prin observarea unui număr suficient de meioze la microscop.
Rata de potrivire este rata la care orice două caractere asociate în generația parentală se recombină diferit în F2.
Dacă cele două loci sunt absolut contigue, probabilitatea ca o chiasmă să le separe va fi practic nulă. Rata de recombinare va fi: n ° recombinanti. Dacă doi loci sunt pe doi cromozomi diferiți, rata de recombinare va fi 0,5 (probabilitate egală, pentru două caractere care au fost unite în generația P, pentru a se găsi aleatoriu împreună în F2). Prin urmare, rata de recombinare poate varia între 0,0 și 0,5. Pentru distanțe mici pe cromozom, distanța și rata de recombinare sunt direct proporționale. Pentru distanțe mai mari există posibilitatea ca două schimburi să aibă loc între doi loci. Acum va apărea clar că doi factori separați de două schimburi sunt din nou asociați. este clar, în acest moment, că se pierde proporționalitatea dintre distanța loci și probabilitatea de recombinare.
Loci găsiți asociați pe același cromozom constituie „grupuri de asociere”. Loci foarte îndepărtați pot avea o astfel de probabilitate de separare prin schimb, încât se comportă ca independenți, dar fiecare dintre aceștia va fi asociat, cu o rată mai mică de recombinare, la locii intermediari.
Când se cunosc ratele de recombinare între multe perechi de gene dintr-un grup de asociere, poate începe construcția „hărților genetice”. Ținând cont de faptul că distanța dintre două gene (a și b) este exprimată prin rata de recombinare și că distanța a de la o a treia genă c poate fi suma sau diferența față de distanța sa de b, este posibil pentru a reconstrui o hartă a distanțelor reciproce, care va fi harta genetică în cadrul acelui grup de asociere, adică al acelui cromozom.
Acum trebuie să luăm în considerare, în general, câteva concepte care limitează manifestarea fenotipică a caracterelor genotipice.
În primul rând vom vorbi despre conceptele de penetranță și expresivitate și apoi vom dedica o atenție deosebită fenomenelor de reglare a acțiunii genetice.
PENETRANȚĂ
Penetrarea unei gene reprezintă capacitatea sa de a se manifesta în fenotip. Penetrarea este măsurată statistic prin numărarea frecvenței fenotipurilor care arată acel caracter din 100 de genotipuri care îl conțin. O trăsătură cu 0,7 penetranță este o trăsătură care apare fenotipic în 70% din frecvența sa genotipică.
EXPRESIVITATE
Expresivitatea este o evaluare cantitativă a gradului de manifestare fenotipică.
REGLEMENTAREA ACȚIUNII GENICE
Celulele produc toate enzimele și proteinele lor cu aceeași viteză și în același timp. Celulele Escherichia coli, de exemplu, pot fi alimentate cu energie și atomi de carbon din lactaza dizaharidă, deoarece acestea sunt capabile să le descompună în glucoză și galactoză datorită enzimei beta-galactozidazei. Într-o E. coli normală care poate avea lactoză, există aproximativ 3 000 de molecule de beta-galactozidază, egale cu 3% din proteinele acelei celule; în absența lactozei va exista o singură moleculă de beta-galactozidază pe celulă bacteriană. Galactozidaza va fi sintetizată din noi molecule de ARNm atunci când poate fi utilizată. Tulpinile mutante de E. coli bogate în enzimă sunt cunoscute chiar și atunci când lactoza este absentă: acești mutanți sunt dezavantajați în comparație cu celulele normale, deoarece sunt forțați la un consum inutil de energie și materiale pentru a produce enzima care va rămâne fără substrat. Substanțele care determină o creștere a cantității de enzime, așa cum este cazul lactozei, vor fi numite inductori, în timp ce enzimele se vor spune că sunt inductibile. Alte substanțe induc, de asemenea, într-un mod specific, producerea anumitor enzime. De asemenea, în E. coli, de exemplu, capabil să-și construiască toți aminoacizii, având carbon și amoniu (NH3), prezența în mediul de cultură a unui anumit aminoacid (histidină, de exemplu) blochează producția tuturor enzimelor asociate cu biosinteza aminoacidului în sine: se va spune despre aceste enzime că sunt represibile. În celulele bacteriene moleculele de ARNm sunt demolate la scurt timp după formarea lor și de aceea controlul producției de ARNm înseamnă controlul sintezei enzimatice în același timp timpul.
OPERONUL
Pentru a explica modul în care celula bacteriană este capabilă să-și controleze propria producție de enzime Jacob și Monod au formulat ipoteza operonului; operonul este format din mai multe gene care sunt funcțional legate și aliniate fără discontinuitate de-a lungul unei întinderi de ADN. Operonul este format din trei tipuri diferite de gene: promotorul, de unde începe formarea ARNm; operatorul, unde se exercită controlul; una sau mai multe gene structurale, care codifică enzimele sau alte proteine. În sistemul beta-galactozidazei, operonul include, pe lângă cel pentru beta-galactozidază, și alte două gene structurale care codifică alte enzime implicate în metabolismul lactozei.Aceste gene sunt adiacente una cu cealaltă și sunt transcrise una după alta de-a lungul aceleiași spirale de ADN într-o singură moleculă de ARNm. Moleculele de ARNm astfel produse sunt active pentru un timp foarte scurt, după care sunt distruse de enzime specifice.
Activitatea operonului este la rândul ei controlată de o altă genă, regulatorul, care poate fi, de asemenea, îndepărtat de operon: acest regulator codifică o proteină, numită represor, care pare să se lege de ADN-ul de la gena operator. Fiind gena operator plasată între promotor și genele structurale blochează de fapt producția de ARNm.
Represorul la rândul său este controlat, iar controlul se efectuează prin intermediul unei substanțe „semnal”. În cazul enzimelor inductibile, această substanță este „inductorul. Inductorul” se leagă de molecula represor modificându-i forma astfel încât să nu se mai poată adapta la ADN: în acest caz, deoarece nu există un represor între promotor și genele structurale , represorul poate forma moleculele de ARNm și din acestea moleculele de proteine. Odată cu epuizarea alimentării cu inductor din nou, regulatorul va recâștiga controlul, ceea ce va opri producția de ARNm nou, deci de proteine noi. În sistemul beta-galactozidazei, inductorul este lactoza sau o substanță foarte similară cu aceasta. se va alătura represorului inactivându-l astfel încât să permită biosinteza enzimelor. În cazul enzimelor represibile, substanța care acționează ca „semnal” acționează ca un corepresor: represorul este activ numai dacă este combinat cu corepresorul. În sistemul histidinei, care implică o duzină de enzime diferite, este acest aminoacid, combinat cu ARNt-ul său, corepresorul.
INTERACȚII ALOSTERICE
Interacțiunile alosterice, care implică inactivarea unei enzime prin modificarea formei acesteia, oferă un mod diferit de reglare a activității metabolice a unei celule. Interacțiunile alosterice permit un control mai precis decât sistemul inductor-represor al operonului, dar nu obțin rezultatul util al excluderii biosintezei unei substanțe date din prima etapă - producerea unui ARNm.
SISTEME DE CONTROL ÎN EUCARIUS
Există câteva fapte care fac să se creadă că un sistem de reglare similar cu operonul funcționează și este preeminent în rândul plantelor și animalelor. celulele sunt foarte diferite.Mecanismul mitozei este de așa natură încât fiecare celulă a unei plante sau animale date posedă toate informațiile
genetică prezentă în ovulul fertilizat. Prin urmare, majoritatea genelor din orice celulă specializată vor rămâne ineficiente pe tot parcursul vieții celulei. ADN-ul din aceste celule este întotdeauna asociat cu proteine. Deci este posibil ca reprimarea genelor în eucariote să necesite tocmai această asociere între ADN și proteine.